|
Переваривание
углеводов
Углеводы, из
которых человек получает энергию, поступают в организм вместе с пищей
в виде сложных форм, таких как ди- и полисахариды (крахмал и гликоген).
Также потребляется целлюлоза, но она не переваривается.
Первый шаг в метаболизме углеводов - это превращение полимеров в мономеры,
которые могут транспортироваться через стенку кишечника.
Расщепление полимеров начинается в ротовой полосте. Слюна имеет слабокислую
рН (6,8) и содержит амилазу, которая начинает переваривание углеводов.
Главным ферментом тонкого кишечника, который расщепляет углеводы,
является альфа-амилаза. Этот фермент секретируется поджелудочной железой
и превращает полисахариды в ди- и трисахариды. Последние превращаются
в моносахариды посредством кишечных сахаридаз (мальтаза, сахараза,
лактаза).
Образовавшееся глюкоза и другие моносахариды транспортируются через
кишичную стенку в v.portae и затем к печёночным гепатоцитам и другим
тканям. Здесь они превращаются в ЖК, аминокислоты, гликоген или окисляются
в клетках.
Окисление глюкозы известно как гликолиз. Глюкоза окисляется либо до
лактата, либо до ПВК. В аэробных условиях ведущим продуктом является
пируват и путь называется аэробный гликолиз.
Когда запасы кислорода исчерпаны, например, во время длительных энергетических
упражнений, ведущим гликолитическим продуктом является лактат (молочная
кислота), и процесс известен как анаэробный гликолиз.
Энергия, получаемая при окислении глюкозы
Аэробный гликолиз глюкозы в пируват требует два моля АТФ с последующим
высвобождением 4АТФ и двух молекул НАДН. Таким образом, превращение
одного моля глюкозы в два моя пирувата сопровождается продукцией двух
молей АТФ и двух молей НАДН.
глюкоза + 2АДФ + 2НАД+ + 2Ф = 2пируват + 2АТФ + 2НАДН + 2Н+
НАДН, получаемый во время гликолиза, используется при синтезе АТФ
через окислительное фосфорилирование, образуя 2-3 моля АТФ в зависимости
от того, какой цикл используется для транспорта электронов от цитоплазматического
НАДН в митохондрию: либо глицеролфосфатный путь, либо малат-аспартатный
путь.
Следовательно, при окисление глюкозы до пирувата получается или 6
или 8 молекул АТФ. Окисление двух молекул пирувата в цикле Кребса
даёт дополнительных 30 молей АТФ. В конечном счёте при окислении 1
моля глюкозы до СО2 и Н2О образуется 36-38 молекул АТФ.
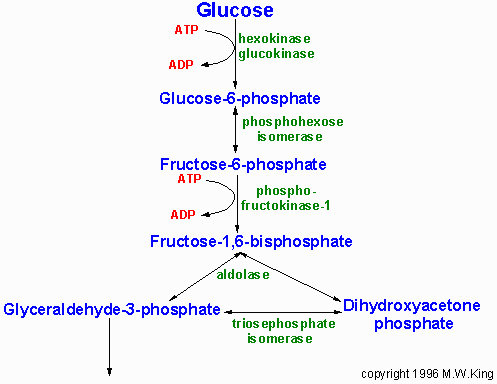
Реакции гликолиза
Гликолиз можно подразделить на два этапа. На первом этапе используется
2 моля АТФ для превращения глюкозы во фруктозо-1,6-дифосфат (Фр-1,6-ФФ).
На втором этапе Фр-1,6-ФФ расщепляется до пирувата с продукцией 4
молей АТФ и 2 молей НАДН.

Гексокиназная
реакция.
Первая реакция гликолиза - превращение глюкозы в глюкозо-6-фосфат
- требует АТФ и катализируется тканеспецифическими изоферментами,
которые называются гексокиназами.
Фосфорилирование имеет две цели:
-гексокиназная реакция превращает неионную глюкозу в анион, который
не может выйти из клетки, т.к. нет транспортных систем для фосфорилированных
сахаров.
-происходит активация глюкозы.
Известно 4 изофермента гексокиназ, тип 4 ещё называется глюкокиназой.
Она обнаружена в печени. Высокая Км глюкокиназы для глюкозы означает,
что этот фермент работает только при высоких концентрациях субстрата.
Эта особенность печёночной глюкокиназы позволяет поддерживать уровень
глюкозы крови. После еды, когда уровень глыкозы достаточно высок,
то происходит активация глюкокиназы. Когда уровень глюкозы низкий,
то такие ткани, как печень и почки (которые содержат глюкокиназу,
но не очень зависимы от глюкозы) не используют её из крови. В то же
время такие ткани как мозг, который зависит от глюкозы продолжает
использовать глюкозу крови, используя гексокиназу с низкой Км для
глюкозы. При недостаточности глюкозы в печени стимулируется глюконеогенез.
Регуляция активности глюкокиназы и гексокиназы также различна. Гексокиназы
1,2 и 3 аллостерически ингибируются продуктом реакции (гл-6-ф), в
то время как глюкокиназа - нет.
Гексозофосфатизомераза.
Вторая реакция гликолиза - это изомеризация, в которой гл-6-ф превращается
в фр-6-ф. Фермент, который катализирует эту реакцию, называется гексозофосфатизомераза
(известна также под названием глюкозофосфатизомераза). Эта реакция
обратима.
Фосфофруктокиназа-1 (ФФрК-1).
Следующая реакция - превращение фр-6-ф в фр-1,6-фф - требует второго
моля АТФ. Эта реакция катализируется фосфофруктокиназой-1. Эта реакция
необратима, поэтому при глюконеогенезе используется фермент фр-1,6-дифосфатаза.
Активность этих двух ферментов тщательно регулируется.
Альдолаза.
Альдолаза катализирует гидролиз фр-1,6-фф на два трёхуглеродных продукта
- фосфодиоксиацетон (ФДА) и глицеральдегид-3-фосфат (3-ФГА). Реакция
идёт в двух направлениях и используется в гликолизе и глюконеогенезе.
Тризофосфатизомераза.
Два продукта альдолазной реакции уравновешены с помощью реакции, которая
катализируется тризофосфатизомеразой. Следующие реакции гликолиза
используют 3-ФГА.
ГлицеральдегидфосфатДГ.
ГлицеральдегидфосфатДГ катализирует НАД+-зависимое окисление 3-ФГА
в 1,3-дифосфоглицерат и НАДН. Эта реакция обратима, и этот же фермент
используется при глюконеогенезе.
Фосфоглицераткиназа.
Высокоэнергетический 1,3-ДФГ используется для образования АТФ и 3-фосфоглицерата
посредством фермента фосфоглицераткиназы. Замечено, что это единственная
реакция гликолиза и глюконеогенеза, которая использует АТФ, но в то
же время обратима.
В эритроцитах с помощью фермента дифосфоглицератмутазы 1,3-ДФГ превращается
в 2,3-ДФГ. Эта реакция очень важна в эритроцитах, т.к. 2,3-ДФГ является
главным регулятором сродства гемоглобина к кислороду. 2,3-ДФГ может
превращаться в 3-фосфоглицерат, который является промежуточным продуктом
гликолиза.
Мутаза и енолаза.
3-ФГ с помощью мутазы превращается в 2-ФГ, а 2-ФГ с помощью енолазы
превращается в фосфоенолпируват (ФЕПВ).
Пируваткиназа.
Последняя реакция гликолиза (аэробного) катализируется высокорегулируемым
ферментом пируваткиназой. Это экзергоническая реакция, т.к. образуется
АТФ.
Анаэробный гликолиз
В аэробных условиях пируват большинства клеток дальше идёт в цикл
Кребса. В анаэробных условиях (а в эритроцитах и в аэробных условиях)
пируват превращается в лактат с помощью фермента лактатдегидрогеназы
(ЛДГ). После этого лактат выходит из клеток в кровь. Реакция превращает
НАДН (который образовался в 3-ФГАДГ-реакции) в НАД+.
Регуляция гликолиза
Ключевые ферменты:
1).Гексокиназа аллостерически ингибируется гл-6-фосфатом. Синтез глюкокиназы
индуцируется инсулином.
2).Фосфофруктокиназа. Ситез индуцируется инсулином. Аллостерические
активаторы - АМФ, фруктозо-2,6-дифосфат, ингибиторы - АТФ, цитрат.
3).Пирувтакиназа. Ситез индуцируется инсулином. Активатор - Фр-1,6-фф,
ингибиторы - АТФ, аланин, ацетилКоА.
Метаболическая судьба пирувата
Пируват является продуктом аэробного гликолиза. Дальнейшая судьба
пирувата зависит от окислительного состояния клетки. В реакции, катализируемой
3-ФГАДГ, моль НАД+ восстанавливается до НАДН. Для того, чтобы поддерживать
редокс-состояние клетки, этот НАДН должен снова окислеться до НАД+.
В аэробном гликолизе это осуществляется в транспорте электронов в
митохондрии, генерируя АТФ. В фосфоглицераткиназной и пируваткиназной
реакциях тоже образуется АТФ, такой путь получения АТФ называется
субстратное фосфорилирование, в то время, как получении энергии при
окислении НАДН называется окислительным фосфорилированием.
В аэробных условиях пируват идёт в цикл Кребса. Пируват вступает в
цикл Кребса в форме ацетилКоА, который образуется в пируватдегидрогеназной
реакции.
В анаэробных условиях пируват превращается в лактат.
Метаболизм лактата
Лактат образуется из пирувата в ходе гликолиза в анаэробных условиях
благодаря действию фермента лактатДГ. Эта реакция обратима. Потом
лактат выходит из клеток и идёт в высокоаэробные ткани, такие как
печень и сердце. В этих тканях лактат с помощью ЛДГ снова окисляется
до пирувата, а пируват идёт в цикл Кребса с получением энергии. Если
в этих тканях энергии итак некуда девать, то пируват используется
как субстрат для синтеза глюкозы (глюконеогенеза).
ЛДГ имеет две различные субъединицы - М и Н. Комбинация этих субъедениц
образует изоферменты ЛДГ, которые имеют разные характеристики. Субъединица
Н превалирует в аэробных тканях, таких как сердечная мышца (Н4-тетрамер),
в то время, как субъединица М доминирует в анаэробных тканях, таких
как скелетная мышца (М4-тетрамер). ЛДГ Н4 имеет низкую Км для пирувата
и ингибируется пируватом, поэтому в сердце никогда не образуется свой
собственный лактат. ЛДГ М4 имеет высокую Км для пирувата и не ингибируется
пируватом.
Метаболизм этанола
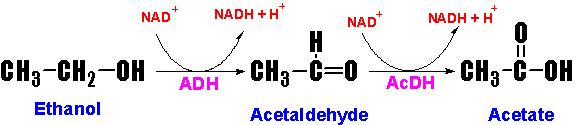
В клетках имеется
фермент алкогольдегидрогеназа (АДГ), который окисляет этанол до ацетальдегида.
Потом ецетальдегид окисляется до ацетата с помощью ацетальдегиддегидрогеназы
(АцДГ). Ацетальдегид и ацетат очень токсичны и ведут к очень многим
побочным эффектам. АДГ и АцДГ катализирует реакцию, ведущую к восстановлении
НАД+ до НАДН.
назад
|


