|
Глюконеогенез
Глюконеогенез - синтез глюкозы из неуглеводных предшественников. Глюконеогенез
необходим в мозге, яичках, эритроцитах и мозговом веществе почек,
где глюкоза является единственным источником энергии. Однако, во время
голодания мозг может получать энергию из кетоновых тел, которые превращаются
в ацетилКоА.
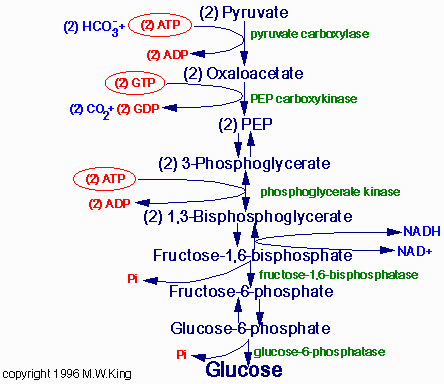
Первоначально
пируват под влиянием пируваткарбоксилазы и при участии СО2 и АТФ карбоксилируется
с образованием оксалоацетата (ЩУК).
Пируваткарбоксилаза находится в митохондриях. Мембрана митохондрий
непроницаема для образовавшегося ЩУКа. Последний здесь же в митохондриях
восстанавливается в малат.
Реакция протекает при участии митохондриальной НАД-зависимой малатдегидрогеназы.
В митохондриях отношение НАДН2/НАД+ относительно высоко, и поэтому
внутримитохондриальный ЩУК легко восстанавливается в малат, который
легко выходит из митохондрии, проходя мембрану митохондрий. В цитоплазме
же отношение НАДН2/НАД+ очень мало, и малат вновь окисляется в ЩУК
при участии НАД-зависимой цитоплазматической малатдегидрогеназы.
Субстраты глюконеогенеза
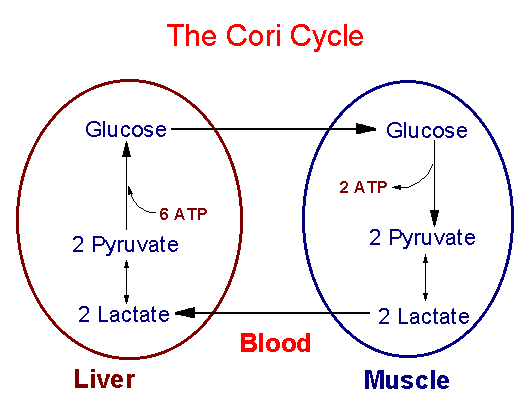
Лактат - образуется в процессе анаэробного гликолиза в эритроцитах
и клетках скелетных мышц. Превращение лактата в глюкозу происходитв
результате работы цикла Кори. Значение - с участием цикла Кори конечные
продукты гликолиза из эритроцитов и скелетных мышц транспортируется
в печень и используются на синтез глюкозы.
Аланин - основная
глюкогенная аминокислота. Превращение аланина в глюкозу происходит
в аланиновом цикле. В скелетных мышцах пируват, образующий в ходе
гликолиза, может превращаться в аланин. Аланин, образующийся в этих
реакциях, является транспортной формой NH2-групп из мышц в печень,
где они в конечном счёте включаются в молекулы мочевины и экскретируются.
При поступлении аланина в гепатоциты он может превращаться в пируват
и использоваться как субстрат в глюконеогенезе. Значение - Аммиак
является чрезвычайно токсическим соединением и основное его количество
обезвреживается в клетках печени (в орнитиновом цикле происходит связывание
его в молекулу мочевины и затем экскреция). Аланин служит транспортной
формой аммиака в печень, где осуществляется его обезвреживание.
Другие аминокислоты. Только две аминокислоты (лейцин и лизин) не могут
использоваться в процессе глюконеогенеза. Это строго кетогенные аминокислоты.
Все остальные глюкогенные аминокислоты при своём метаболизме дают
промежуточные продукты гликолиза или цикла Кребса.
Глицерол. Образуется при катаболизме триацилглицеролов в клетках жировой
ткани, выходит в кровь и затем попадает в печень, где под действием
двух ферментов (глицеролкиназа и альфа-глицеролфосфатдегидрогеназа)
превращается в фосфодиоксиацетон (ФДА), промежуточный продукт гликолиза.
Пропионил-КоА. Бетта-окисление ЖК с нечётным числом атомов углерода
и метаболизм некоторых аминокислот (валин, изолейцин, триптофан, метионин)
сопровождается образованием пропионил-КоА, который может быть превращён
в сукцинил-КоА (метаболит цикла Кребса) под действием двух ферментов:
-пропионил-КоА-карбоксилаза (в качестве кофермента используется биотин
- смотри витамин Н).
-метилмалонил-КоА-мутаза (метилкобаламин в качестве кофермента - смотри
витамин В12).
Суммарное
уравнение:
2ПВК + 4АТФ + 2ГТФ + 2НАДН + 2Н+ + 6Н2О = глюкоза + 4АДФ + 2ГДФ +
6Ф + 2НАД+
На синтез молекулы глюкозы из двух молекул пирувата расходуется 4АТФ
и 2ГТФ. Процесс окисления ЖК поставляет энергию для глюконеогенеза.
Для восстановительных этапов требуется две молекулы НАДН.
Пируваткарбоксилаза, катализирующая первую реакцию, имеет аллостерического
активатора - ацетилКоА.
Регуляция глюконеогенеза
1).Репрессируется после приёма богатой углеводами пищи (под действием
инсулина) и индуцируется при голодании, стрессе, диабете (под действием
глюкокортикоидов).
2).Процесс окисления ЖК стимулирует глюконеогенез. Стимуляция осуществляется
через увеличение уровня ацетил-КоА.
3).Реципрокная взаимосвязь:
-ацетилКоА ингибирует пируватДГ и активирует пируваткарбоксилазу.
-АТФ активирует фруктозодифосфатазу, АМФ - ингибирует.
-фруктозо-2,6-дифосфат активирует фосфофруктокиназу-1 и ингибирует
фруктозодифосфатазу-1.
назад
|


